The Dirk Remus Lab
Research

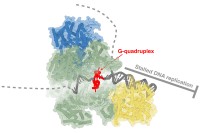
Our lab studies the mechanism of eukaryotic chromosome replication. Chromosomes are the carriers of the genetic and epigenetic information and faithful chromosome replication is of fundamental importance for genome maintenance during normal cell proliferation. We employ fully reconstituted yeast and human DNA replication systems to investigate the structure and biochemical mechanism of the eukaryotic replication machinery. A particular focus of the lab is to elucidate the molecular consequences of replication stress, which encompasses conditions that impede the normal progression of replication forks, and which is a major driver of cancer-associated genome instability. Through this approach, we aim to understand how faithful and efficient chromosomal DNA replication is achieved under normal conditions and, conversely, how the replication machinery is affected by and overcomes the diverse array of physical obstacles encountered on the chromosomal template. Ultimately, our work aims to uncover novel cancer therapeutic targets to control the dysregulated proliferation of cancer cells.

Featured News


People
Dirk Remus, PhD
- Molecular biologist Dirk Remus investigates mechanisms of DNA replication in eukaryotic cells.
- PhD, University of California, Berkeley
- [email protected]
- Email Address
- 212-639-5263
- Office Phone
Members




























Achievements
- Louis V. Gerstner, Jr. Young Investigator Award, Memorial Sloan Kettering Cancer Center (2011)
- Long-Term Fellow, EMBO (2006)