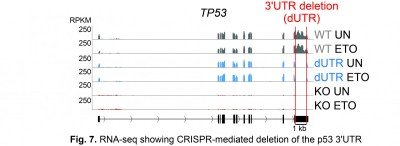
We use CRISPR to delete 3′UTRs at their endogenous loci and study the functional outcomes (Fig. 7).
We use CRISPR to delete enhancers to change 3′UTR ratios of endogenous genes.
We perform functional screens in S. cerevisiae to identify 3′UTR functions that go beyond the regulation of protein abundance.
We are developing new assays to study 3′UTR functions independently of protein abundance regulation.
Quantitative mapping of 3′UTRs in human, mouse, and yeast
We previously established a quantitative tag-based sequencing protocol, called 3′-seq, to map 3′UTR boundaries and 3′UTR isoform usage in tissues and cell lines (Lianoglou, Genes Dev 2013). Our study was chosen by Science Signaling as one of the Signaling Breakthroughs of 2013 (Science Signaling). Following up on this initial study, we examined intronic polyadenylation (IPA) isoform expression in 60 diverse samples, including a large number of normal and malignant immune cell types (Singh, Nat commun 2018) and showed that truncated proteins generated by IPA can inactivate tumor-suppressor genes in leukemia (Lee, Singh, Nature 2018).
We currently use single cell RNA-seq data to identify 3′UTR ratios at cell type resolution (Fig. 8).
We mapped the 3′UTRs of S. cerevisiae across different metabolic conditions (Fig. 9).
Functional and molecular characterization of TIS granules
We purified TIS granules and identified the enriched mRNAs. We are performing mass spectrometry analysis on TIS granules, and we use TIS11B KO cells to assess the functions of TIS granules.
We are using in vivo and in vitro assays to determine the surface properties of proteins that enable their enrichment in TIS granules.
We use genetically encoded multimeric nanoparticles to assess diffusion coefficients in TIS granules.
We are assessing the role of mRNAs in the 3D-organization of TIS granules.
One of our goals is to make designer organelles to promote or inhibit certain reactions.
Cytoplasmic compartmentalization by membraneless organelles
We are identifying new cytoplasmic membraneless organelles that are associated with membranes.
We are identifying reactions that are promoted in membraneless organelles.
3′UTR-mediated co-translational protein complex assembly
We hypothesize that 3′UTRs play major roles in co-translational assembly as they may control spatial proximity of the translated mRNAs. mRNAs that encode subunits of protein complexes may co-localize through bridge proteins that bind to 3′UTR-bound RNA-binding proteins or in cytoplasmic RNA granules (Mayr, Nature N&V 2018).